SMB2021
June 14-17, 2021
Non-Equilibrium Thermodynamics in Biology:
from Chemical Reaction Networks to Natural Selection
Larry Li,
Bill
Cannon and I ran a session non-equilibrium thermodynamics in biology
at SMB2021, the annual meeting of the
Society for Mathematical Biology. You can see talk slides here!
Here is the basic idea:
Since Lotka, physical scientists have argued that living things belong
to a class of complex and orderly systems that exist not despite the
second law of thermodynamics, but because of it. Life and evolution,
through natural selection of dissipative structures, are based on
non-equilibrium thermodynamics. The challenge is to develop an
understanding of what the respective physical laws can tell us about
flows of energy and matter in living systems, and about growth, death
and selection. This session will address current challenges including
understanding emergence, regulation and control across scales, and
entropy production, from metabolism in microbes to evolving
ecosystems.
Click on the links to see talk slides:
-
Persistence, permanence,
and global stability in reaction network models: some results inspired
by thermodynamic principles
Gheorghe Craciun, University of Wisconsin–Madison
The standard mathematical model for the dynamics of concentrations in
biochemical networks is called mass-action kinetics. We describe
mass-action kinetics and discuss the connection between special
classes of mass-action systems (such as detailed balanced and complex
balanced systems) and the Boltzmann equation. We also discuss the
connection between the 'global attractor conjecture' for complex
balanced mass-action systems and Boltzmann's H-theorem. We also
describe some implications for biochemical mechanisms that implement
noise filtering and cellular homeostasis.
-
The
principle of maximum caliber of nonequilibria
Ken Dill, Stony Brook University
Maximum Caliber is a principle for inferring pathways and rate
distributions of kinetic processes. The structure and foundations of
MaxCal are much like those of Maximum Entropy for static
distributions. We have explored how MaxCal may serve as a general
variational principle for nonequilibrium statistical physics - giving
well-known results, such as the Green-Kubo relations, Onsager's
reciprocal relations and Prigogine's Minimum Entropy Production
principle near equilibrium, but is also applicable far from
equilibrium. I will also discuss some applications, such as finding
reaction coordinates in molecular simulations non-linear dynamics in
gene circuits, power-law-tail distributions in 'social-physics'
networks, and others.
-
Nonequilibrium biomolecular
information processes
Pierre Gaspard, Université libre de Bruxelles
Nearly 70 years have passed since the discovery of DNA
structure and its role in coding genetic information. Yet, the
kinetics and thermodynamics of genetic information processing in DNA
replication, transcription, and translation remain poorly
understood. These template-directed copolymerization processes are
running away from equilibrium, being powered by extracellular energy
sources. Recent advances show that their kinetic equations can be
exactly solved in terms of so-called iterated function
systems. Remarkably, iterated function systems can determine the
effects of genome sequence on replication errors, up to a million
times faster than kinetic Monte Carlo algorithms. With these new
methods, fundamental links can be established between molecular
information processing and the second law of thermodynamics, shedding
a new light on genetic drift, mutations, and evolution.
-
Nonequilibrium dynamics of
disturbed ecosystems
John Harte, University of California, Berkeley
The Maximum Entropy Theory of Ecology (METE) predicts the shapes of
macroecological metrics in relatively static ecosystems, across
spatial scales, taxonomic categories, and habitats, using constraints
imposed by static state variables. In disturbed ecosystems, however,
with time-varying state variables, its predictions often fail. We
extend macroecological theory from static to dynamic, by combining the
MaxEnt inference procedure with explicit mechanisms governing
disturbance. In the static limit, the resulting theory, DynaMETE,
reduces to METE but also predicts a new scaling relationship among
static state variables. Under disturbances, expressed as shifts in
demographic, ontogenic growth, or migration rates, DynaMETE predicts
the time trajectories of the state variables as well as the
time-varying shapes of macroecological metrics such as the species
abundance distribution and the distribution of metabolic rates over
individuals. An iterative procedure for solving the dynamic theory is
presented. Characteristic signatures of the deviation from static
predictions of macroecological patterns are shown to result from
different kinds of disturbance. By combining MaxEnt inference with
explicit dynamical mechanisms of disturbance, DynaMETE is a candidate
theory of macroecology for ecosystems responding to anthropogenic or
natural disturbances.
-
Stochastic chemical
reaction networks
Supriya Krishnamurthy, Stockholm University
The study of chemical reaction networks (CRN's) is a very active
field. Earlier well-known results (Feinberg Chem. Enc. Sci. 42 2229
(1987), Anderson et al Bull. Math. Biol. 72 1947 (2010)) identify a
topological quantity called deficiency, easy to compute for CRNs of
any size, which, when exactly equal to zero, leads to a unique
factorized (non-equilibrium) steady-state for these networks. No
general results exist however for the steady states of
non-zero-deficiency networks. In recent work, we show how to write the
full moment-hierarchy for any non-zero-deficiency CRN obeying
mass-action kinetics, in terms of equations for the factorial
moments. Using these, we can recursively predict values for lower
moments from higher moments, reversing the procedure usually used to
solve moment hierarchies. We show, for non-trivial examples, that in
this manner we can predict any moment of interest, for CRN's with
non-zero deficiency and non-factorizable steady states. It is however
an open question how scalable these techniques are for large networks.
-
Heat flows adjust local ion concentrations in favor of prebiotic chemistry
Christof Mast, Ludwig-Maximilians-Universität München
Prebiotic reactions often require certain initial
concentrations of ions. For example, the activity of RNA enzymes
requires a lot of divalent magnesium salt, whereas too much monovalent
sodium salt leads to a reduction in enzyme function. However, it is
known from leaching experiments that prebiotically relevant
geomaterial such as basalt releases mainly a lot of sodium and only
little magnesium. A natural solution to this problem is heat fluxes
through thin rock fractures, through which magnesium is actively
enriched and sodium is depleted by thermogravitational convection and
thermophoresis. This process establishes suitable conditions for
ribozyme function from a basaltic leach. It can take place in a
spatially distributed system of rock cracks and is therefore
particularly stable to natural fluctuations and disturbances.
-
Deficiency of chemical reaction networks and
thermodynamics
Matteo Polettini, University of Luxembourg
Deficiency is a topological property of a Chemical Reaction Network
linked to important dynamical features, in particular of deterministic
fixed points and of stochastic stationary states. Here we link it to
thermodynamics: in particular we discuss the validity of a strong
vs. weak zeroth law, the existence of time-reversed mass-action
kinetics, and the possibility to formulate marginal fluctuation
relations. Finally we illustrate some subtleties of the Python module
we created for MCMC stochastic simulation of CRNs, soon to be made
public.
-
Large deviations theory
and emergent landscapes in biological dynamics
Hong Qian, University of Washington
The mathematical theory of large deviations provides a nonequilibrium
thermodynamic description of complex biological systems that consist
of heterogeneous individuals. In terms of the notions of stochastic
elementary reactions and pure kinetic species, the continuous-time,
integer-valued Markov process dictates a thermodynamic structure that
generalizes (i) Gibbs’ macroscopic chemical thermodynamics of
equilibrium matters to nonequilibrium small systems such as living
cells and tissues; and (ii) Gibbs’ potential function to the
landscapes for biological dynamics, such as that of C. H. Waddington’s
and S. Wright’s.
-
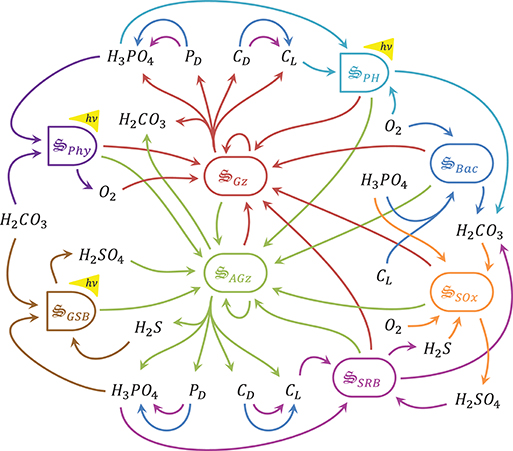
Using the maximum entropy production
principle to understand and predict microbial biogeochemistry
Joseph Vallino, Marine Biological Laboratory, Woods Hole
Natural microbial communities contain billions of individuals per
liter and can exceed a trillion cells per liter in sediments, as well
as harbor thousands of species in the same volume. The high species
diversity contributes to extensive metabolic functional capabilities
to extract chemical energy from the environment, such as
methanogenesis, sulfate reduction, anaerobic photosynthesis,
chemoautotrophy, and many others, most of which are only expressed by
bacteria and archaea. Reductionist modeling of natural communities is
problematic, as we lack knowledge on growth kinetics for most
organisms and have even less understanding on the mechanisms governing
predation, viral lysis, and predator avoidance in these systems. As a
result, existing models that describe microbial communities contain
dozens to hundreds of parameters, and state variables are extensively
aggregated. Overall, the models are little more than non-linear
parameter fitting exercises that have limited, to no, extrapolation
potential, as there are few principles governing organization and
function of complex self-assembling systems. Over the last decade, we
have been developing a systems approach that models microbial
communities as a distributed metabolic network that focuses on
metabolic function rather than describing individuals or species. We
use an optimization approach to determine which metabolic functions in
the network should be up regulated versus those that should be down
regulated based on the non-equilibrium thermodynamics principle of
maximum entropy production (MEP). Derived from statistical mechanics,
MEP proposes that steady state systems will likely organize to
maximize free energy dissipation rate. We have extended this
conjecture to apply to non-steady state systems and have proposed that
living systems maximize entropy production integrated over time and
space, while non-living systems maximize instantaneous entropy
production. Our presentation will provide a brief overview of the
theory and approach, as well as present several examples of applying
MEP to describe the biogeochemistry of microbial systems in laboratory
experiments and natural ecosystems.
-
Reduction and the
quasi-steady state approximation
Carsten Wiuf, University of Copenhagen
Chemical reactions often occur at different time-scales. In
applications of chemical reaction network theory it is often desirable
to reduce a reaction network to a smaller reaction network by
elimination of fast species or fast reactions. There exist various
techniques for doing so, e.g. the Quasi-Steady-State Approximation or
the Rapid Equilibrium Approximation. However, these methods are not
always mathematically justifiable. Here, a method is presented for
which (so-called) non-interacting species are eliminated by means of
QSSA. It is argued that this method is mathematically sound. Various
examples are given (Michaelis-Menten mechanism, two-substrate
mechanism, ...) and older related techniques from the 50-60ies are
briefly discussed.
Copyright held by the authors; figure by Joseph Vallino and Julie Huber.
baez@math.removethis.ucr.andthis.edu